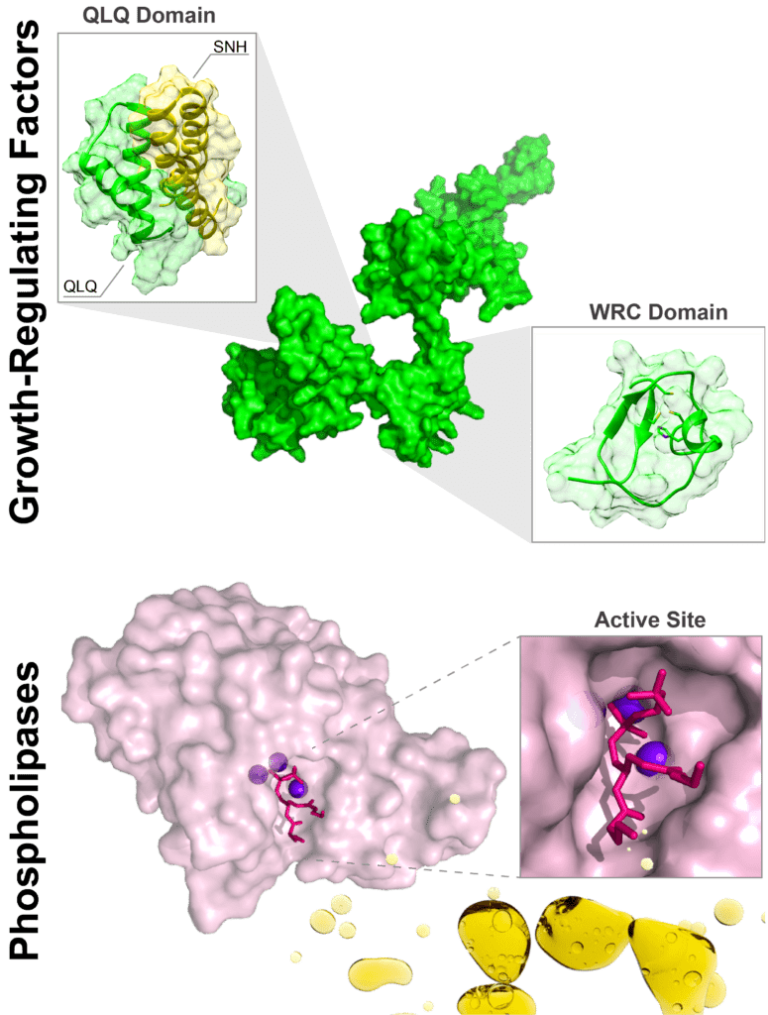
Characterization of a novel thermostable phospholipase C from T. kodakarensis suitable for oil degumming.
Appl Microbiol Biotechnol. Epub ahead of print. Marchisio F, Di Nardo L, Val DS, Cerminati S, Espariz M, Rasia RM, Menzella HG, Castelli ME. (2022).